")
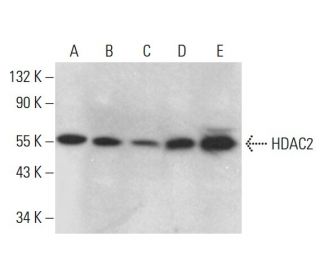
Histone Deacetylase 2 (HDAC2) Antibody (F-6): sc-55542
- Histone Deacetylase 2 (HDAC2) Antibody (F-6) is a mouse monoclonal IgG1 κ, cited in 3 publications, provided at 200 µg/ml
- recommended for detection of HDAC2 of mouse, rat and human origin by WB, IP, IF and ELISA
- See Histone Deacetylase 2 (C-8): sc-9959 for Histone Deacetylase 2 (HDAC2) antibody conjugates, including AC, HRP, FITC, PE, Alexa Fluor® 488, 594, 647, 680 and 790.
- m-IgG Fc BP-HRP, m-IgG1 BP-HRP and m-IgGκ BP-HRP are the preferred secondary detection reagents for Histone Deacetylase 2 (HDAC2) Antibody (F-6) for WB applications. These reagents are now offered in bundles with Histone Deacetylase 2 (HDAC2) Antibody (F-6) (see ordering information below).

HDAC2 Antibody (F-6) is a mouse monoclonal IgG1 antibody that detects HDAC2 in mouse, rat, and human samples through applications such as western blotting (WB), immunoprecipitation (IP), immunofluorescence (IF), and enzyme-linked immunosorbent assay (ELISA). HDAC2, or Histone Deacetylase 2, plays a crucial role in the regulation of gene expression by removing acetyl groups from histones, leading to chromatin condensation and transcriptional repression. This function is vital for maintaining cellular homeostasis and regulating processes such as cell cycle progression, differentiation, and apoptosis. Dysregulation of HDAC2 has been implicated in various diseases, including cancer, where HDAC2 overexpression can lead to silencing of tumor suppressor genes. Additionally, HDAC2 interacts with several key proteins, including the transcriptional co-repressor Sin3A and the NuRD complex, which further underscores HDAC2′s importance in the regulation of gene expression and chromatin dynamics. Anti-HDAC2 antibody (F-6) effectively targets HDAC2, making this reagent an invaluable tool for researchers studying epigenetic modifications and their implications in health and disease.
Ordering Information
| Product Name | Catalog # | UNIT | Price | Qty | FAVORITES | |
Histone Deacetylase 2 (HDAC2) Antibody (F-6) | sc-55542 | 200 µg/ml | $322.00 | |||
Histone Deacetylase 2 (HDAC2) Antibody (F-6): m-IgG Fc BP-HRP Bundle | sc-537069 | 200 µg Ab; 10 µg BP | $361.00 | |||
Histone Deacetylase 2 (HDAC2) Antibody (F-6): m-IgGκ BP-HRP Bundle | sc-534255 | 200 µg Ab; 40 µg BP | $361.00 | |||
Histone Deacetylase 2 (HDAC2) Antibody (F-6): m-IgG1 BP-HRP Bundle | sc-544986 | 200 µg Ab; 20 µg BP | $361.00 |