")
ATP13A2 Plasmídeo de ativação de CRISPR (h): sc-404770-ACT
- alvos específicos: human
- 20 µg de plasmídeo de DNA pronto para transfecção; Suficiente para até 20 transfecções
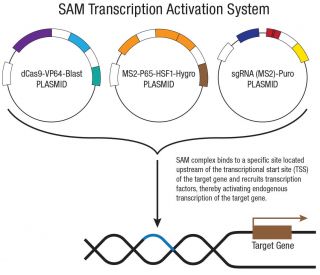
- ATP13A2O plasmídeo de ativação de CRISPR (h)e um mediador da ativação sinergética (SAM) dentro do sistema de ativada da transcrição, criado para a especificamente fazer a regulação genética crescente
- ATP13A2 Plasmídeo de ativação CRISPR (h) consiste em 3 pares de plasmídeos com a razão de massa de 1:1:1: um plasmídeo contento o código para Cas9 desativada
- O complexo SAM resultante se liga a uma região especifica a qual contem aproximadamente 200-250 nt na região upstream da região de inicio da transcrição e fornece um recrutamento robusto de fatores de transcrição para uma eficiente ativação genética.
- Os gRNAs codificados pelo Plasmídeo de Ativação CRISPR ATP13A2 (h) e pelo Plasmídeo de Ativação CRISPR ATP13A2 (h2) têm como alvo regiões reguladoras distintas a montante do local de início da transcrição de ATP13A2. Um ou ambos os desenhos podem estar disponíveis
- Após a transfecção, a eficácia do processo de nocaute genético por ser testada WB, IF ou IHC usando o anticorpo:ATP13A2 Anticorpo (4B7): sc-293367

Informacoes sobre ordens
| Nome do Produto | Numero de Catalogo | UNID | Preco | Qde | FAVORITOS | |
ATP13A2 Plasmídeo de ativação de CRISPR (h) | sc-404770-ACT | 20 µg | $397.00 |
ATP13A2 codifica uma ATPase lisossomal do tipo P5, implicada na manutenção da homeostase endolisossomal, incluindo a função da membrana lisossomal, o manejo de cátions e poliaminas e o tráfego vesicular. Ao sustentar o controle de qualidade dependente de lisossomos, a ATP13A2 influencia o fluxo autofagia–lisossomo e vias de proteostase que se cruzam com respostas ao estresse mitocondrial e ao dano oxidativo. A disrupção ou desregulação de ATP13A2 tem sido associada a fenótipos neurodegenerativos, com associações recorrentes ao parkinsonismo de início juvenil e a defeitos semelhantes a doenças de depósito lisossomal observados em modelos celulares. Como resultado, a ATP13A2 é amplamente estudada por seu papel na vulnerabilidade neuronal, na depuração de proteínas propensas à agregação e em redes de sinalização endolisossomal.
ATP13A2 O Plasmídeo de Ativação CRISPR (h) oferece uma abordagem direcionada e não destrutiva para regular positivamente a expressão endógena de ATP13A2 sem alterar a sequência de ADN subjacente.
ATP13A2 O Plasmídeo de Ativação CRISPR (h) é um sistema mediador de ativação sinérgica (SAM) de três plasmídeos, concebido para a regulação positiva transcricional altamente eficiente e específica do locus ATP13A2 em linhas celulares humanas. O sistema é construído em torno de uma Cas9 cataliticamente inativa (dCas9) portadora de duas mutações inativadoras (D10A e N863A) que eliminam a atividade nuclease, preservando simultaneamente a ligação ao ADN. Esta dCas9 é fundida com VP64, um potente ativador transcricional, e é coexpressa com um gene de resistência à blasticidina para seleção. O segundo plasmídeo codifica a proteína de fusão MS2-p65-HSF1, um complexo ativador secundário que atua em conjunto com o dCas9-VP64, juntamente com um gene de resistência à higromicina. O terceiro plasmídeo codifica um sgRNA de 20 nt específico para o alvo, fundido a dois aptâmeros de RNA MS2 que recrutam o complexo MS2-p65-HSF1 para o local de ativação, acompanhado por um gene de resistência à puromicina. Os três plasmídeos são administrados numa proporção de massa de 1:1:1 para uma expressão equilibrada de todos os componentes do sistema.
Uma vez montado no locus alvo, o complexo SAM liga-se a cerca de 200 pb a montante do local de início da transcrição ATP13A2, onde VP64, p65 e HSF1 atuam em conjunto para recrutar a maquinaria transcricional e impulsionar a regulação positiva da expressão endógena de ATP13A2. Ao contrário da Cas9 com atividade nuclease, o dCas9 não introduz quebras de cadeia dupla nem modifica a sequência genómica, preservando o locus ATP13A2 nativo e permitindo o estudo de respostas transcricionais dependentes de ATP13A2 no locus endógeno, tornando-o uma ferramenta valiosa para estudos funcionais, identificação de genes-alvo e modelagem da restauração da via ATP13A2 em células tumorais com expressão de ATP13A2 silenciada ou reduzida.
Apenas para uso em investigação. Não se destina a uso diagnóstico ou terapêutico.