")
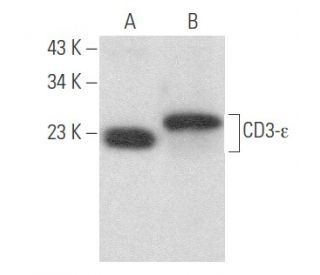
CD3-ε Anticuerpo (G-5): sc-137096
- CD3-ε Anticuerpo (G-5) es un monoclonal de ratón IgG1 κ, ver las 1 publicaciones, proporcionado como 200 µg/ml
- producido contra los amino ácidos 1-207 de la proteina total CD3-ε de origen human
- recomendado para detectarCD3-ε de mouse, rat y human origen, mediante WB, IP, IF, IHC(P) y ELISA
- Ver CD3-ε (UCH-T1): sc-1179 para CD3-ε anticuerpos conjugados, incluyendo AC, HRP, FITC, PE, Alexa Fluor® 488, 594, 647, 680 y 790.
- m-IgG Fc BP-HRP, 1 BP-HRP">m-IgG1 BP-HRP y m-IgGκ BP-HRP son los reactivos de detección secundarios preferidos para CD3-ε Anticuerpo (G-5) para aplicaciones WB e IHC(P). Estos reactivos se ofrecen ahora en paquetes con CD3-ε Anticuerpo (G-5)(véase la información de pedido más abajo).

El receptor de antígeno de células T (TCR) reconoce antígenos extraños y traduce tales eventos de reconocimiento en señales intracelulares que provocan un cambio en la célula de un estado inactivo a un estado activado. Gran parte de este proceso de señalización se puede atribuir a un complejo de proteínas de múltiples subunidades que se asocia directamente con el TCR. Este complejo ha sido designado como CD3 (grupo de diferenciación 3). Está compuesto por cinco cadenas de polipéptidos invariantes que se asocian para formar tres dímeros: un heterodímero de las cadenas gamma y epsilon (CD3-γ y CD3-ε), un heterodímero de las cadenas delta y epsilon (CD3-δ y CD3-ε) y un homodímero de dos cadenas zeta (CD3-ζ) o un heterodímero de las cadenas zeta y eta (CD3-ζ y CD3-η). CD3-ζ y CD3-η están codificados por el mismo gen, pero difieren en sus extremos carboxilo terminales debido a un evento de empalme alternativo. CD3-γ, CD3-ε y CD3-δ cada uno contiene una copia única de un motivo conservado de activación de tirosina basado en inmunorreceptor (ITAM). En contraste, CD3-ζ contiene tres copias consecutivas del mismo motivo. Los ITAMs fosforilados actúan como sitios de acoplamiento para proteínas quinasas como ZAP-70 y Syk y también son capaces de regular su actividad quinasa. La estructura cristalina de los dominios SH2 de ZAP-70 unidos a los ITAMs de CD3-ζ ha sido resuelta.
Información sobre pedidos
| Nombre del producto | Número de catálogo | UNIDAD | Precio | CANTIDAD | Favoritos | |
CD3-ε Anticuerpo (G-5) | sc-137096 | 200 µg/ml | $322.00 | |||
Paquete de CD3-ε (G-5): m-IgG Fc BP-HRP | sc-553218 | 200 µg Ab; 10 µg BP | ||||
Paquete de CD3-ε (G-5): m-IgGκ BP-HRP | sc-553217 | 200 µg Ab; 40 µg BP | ||||
Paquete de CD3-ε (G-5): m-IgG1 BP-HRP | sc-553219 | 200 µg Ab; 20 µg BP |