")
ATP5B CRISPR Activation Plasmid (h): sc-401009-ACT
- Zielspezies: human
- 20 µg transfektionsfertige, aufgereinigte Plasmid DNA; geeignet für 20 Transfektionen
- ATP5B CRISPR Activation Plasmid (h) ist ein Transkriptionsaktivierungs System (SAM) welches für die gezielte Verstärkung der Genexpression bestimmt ist
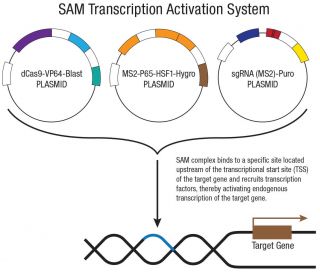
- ATP5B CRISPR Aktivierungsplasmide (h) bestehen aus 3 Plasmiden im Massenverhältnis 1:1:1: ein Plasmid kodiert für die deaktivierte Cas9 (dCas9) Nuklease (D10A und N863A) fusioniert an die Transaktivierungsdomaine VP64 sowie ein Gen für die Blasticidin Resistenz; ein zweites Plasmid kodierend für das MS2-p65-HSF1 Fusionsprotein sowie ein Gen für die Hygromycin Resistenz; ein drittes Plasmid kodierd für die Ziel-spezifische 20 nt guide RNA fusioniert an zwei MS2 RNA Aptamere sowie ein Gen für die Puromycin Resistenz.
- Der entstehende SAM-Komplex (Mediator-Komplex zur synergistischen Gen-Aktivierung) bindet eine sequenzspezifische Region 200-250 nt upstream (in 5'-Richtung) des Transkriptionsstartsignals und rekrutiert dort ständig Transkriptionsfaktoren für eine verstärkte Gen-Aktivierung und Gen-Expression.
- Die vom ATP5B CRISPR-Aktivierungsplasmid (h) und vom ATP5B CRISPR-Aktivierungsplasmid (h2) kodierten gRNAs zielen auf unterschiedliche regulatorische Regionen stromaufwärts der ATP5B-Transkriptionsstartstelle ab. Eines oder beide Designs sind möglicherweise verfügbar
- Nach der Transfektion kann die Effizienz des Gen-Knockouts per Western Blot oder histologisch mit folgendem Antikörper überprüft werden: ATP5B: sc-55597

Bestellinformation
| Produkt | Katalog # | EINHEIT | Preis | ANZAHL | Favoriten | |
ATP5B CRISPR Activation Plasmid (h) | sc-401009-ACT | 20 µg | $397.00 |
ATP5B kodiert die Beta‑Untereinheit der mitochondrialen ATP‑Synthase (Komplex V), eine zentrale katalytische Komponente des F1‑Sektors, die während der oxidativen Phosphorylierung die protonenmotorische Kraft mit der ATP‑Produktion koppelt. Durch die Unterstützung der zellulären Bioenergetik und die Aufrechterhaltung des mitochondrialen Membranpotenzials beeinflusst ATP5B Prozesse wie die Homöostase reaktiver Sauerstoffspezies, die Anfälligkeit für Apoptose und die metabolische Anpassung. Eine veränderte Expression oder Funktion von ATP‑Synthase‑Untereinheiten, einschließlich ATP5B, wurde mit Phänotypen mitochondrialer Dysfunktion in Verbindung gebracht, wie sie bei Neurodegeneration, kardiometabolischem Stress und der krebsassoziierten metabolischen Umprogrammierung beobachtet werden, weshalb es in Studien zur Mitochondrienbiologie häufig als Readout dient. ATP5B wird daher oft in Signal- und Regulationswegen untersucht, die die Aktivität der Elektronentransportkette mit dem ATP‑Bedarf, der mitochondrialen Qualitätskontrolle und Stressantwort‑Signalen verknüpfen.
ATP5B Das CRISPR-Aktivierungsplasmid (h) bietet einen gezielten, nicht-destruktiven Ansatz zur Hochregulierung der endogenen ATP5B-Expression, ohne die zugrunde liegende DNA-Sequenz zu verändern.
ATP5B Das CRISPR-Aktivierungsplasmid (h) ist ein aus drei Plasmiden bestehendes synergistisches Aktivierungsmediator-System (SAM), das für eine hocheffiziente, ortsspezifische transkriptionelle Hochregulation des ATP5B-Lokus in menschlichen Zelllinien entwickelt wurde. Das System basiert auf einem katalytisch inaktiven Cas9 (dCas9), das zwei inaktivierende Mutationen (D10A und N863A) trägt, welche die Nukleaseaktivität eliminieren, während die DNA-Bindung erhalten bleibt. Dieses dCas9 ist mit VP64, einem potenten Transkriptionsaktivator, fusioniert und wird zusammen mit einem Blasticidin-Resistenzgen zur Selektion koexprimiert. Das zweite Plasmid kodiert das MS2-p65-HSF1-Fusionsprotein, einen sekundären Aktivatorkomplex, der zusammen mit dCas9-VP64 wirkt, sowie ein Hygromycin-Resistenzgen. Das dritte Plasmid kodiert für eine zielspezifische 20-nt-sgRNA, die an zwei MS2-RNA-Aptamere fusioniert ist, welche den MS2-p65-HSF1-Komplex an die Aktivierungsstelle rekrutieren, begleitet von einem Puromycin-Resistenzgen. Die drei Plasmide werden im Massenverhältnis 1:1:1 verabreicht, um eine ausgewogene Expression aller Systemkomponenten zu gewährleisten.
Nach der Assemblierung am Zielort bindet der SAM-Komplex etwa 200 bp stromaufwärts der ATP5B-Transkriptionsstartstelle, wo VP64, p65 und HSF1 gemeinsam die Transkriptionsmaschinerie rekrutieren und die Hochregulation der endogenen ATP5B-Expression vorantreiben. Im Gegensatz zu nukleaseaktivem Cas9 verursacht dCas9 keine Doppelstrangbrüche und verändert die genomische Sequenz nicht, wodurch der native ATP5B-Locus erhalten bleibt und die Untersuchung von ATP5B-abhängigen Transkriptionsreaktionen am endogenen Locus ermöglicht wird. Dies macht es zu einem wertvollen Werkzeug für Funktionsstudien, die Identifizierung von Zielgenen und die Modellierung der Wiederherstellung des ATP5B-Signalwegs in Tumorzellen mit stillgelegtem oder reduziertem ATP5B-Ausdruck.
Nur für Forschungszwecke. Nicht für diagnostische oder therapeutische Zwecke bestimmt.