")
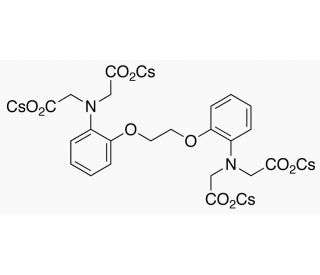

BAPTA-tetracesium Salt (CAS 480436-84-8)
Voir les citations produits (1)
ACCÈS RAPIDE AUX LIENS
Le sel de BAPTA-tétracesium (BAPTA-TC) est un composé largement utilisé dans la recherche scientifique en raison de son large éventail d'applications et de sa capacité à se lier efficacement aux ions calcium. Dérivé du produit chimique BAPTA (acide 1,2-bis(2-aminophénoxy)éthane-N,N,N',N'-tétraacétique), initialement synthétisé dans les années 1970, le sel de BAPTA-tétracesium fonctionne comme un agent chélateur employé pour lier et moduler les concentrations de calcium dans les cellules. La polyvalence du sel de BAPTA-tétracesium est évidente dans son utilisation à diverses fins. Il permet de modifier les concentrations cellulaires de calcium, ce qui facilite l'examen de l'impact du calcium sur les processus cellulaires. En outre, il a été utilisé dans des études de développement de médicaments pour moduler les effets des médicaments sur les cellules. Le sel de BAPTA-tétracesium a également été un outil précieux dans l'étude des effets du calcium sur l'excitabilité neuronale, la contraction musculaire et les voies de signalisation intracellulaires. En se liant aux ions calcium, le sel de BAPTA-tétracesium entrave leur interaction avec d'autres molécules telles que les protéines et les enzymes. Par conséquent, il empêche l'activation des processus cellulaires dépendant du calcium, y compris la contraction musculaire et l'excitabilité neuronale. De plus, la liaison du sel de BAPTA-tétracesium aux ions calcium empêche l'activation des voies de signalisation intracellulaires, ce qui facilite l'étude de l'influence du calcium sur les mécanismes cellulaires.
BAPTA-tetracesium Salt (CAS 480436-84-8) Références
- Régulation de I(Cl,swell) dans les cellules de neuroblastome par les voies de signalisation des protéines G. | Estevez, AY., et al. 2001. Am J Physiol Cell Physiol. 281: C89-98. PMID: 11401830
- Effet du pH extracellulaire sur les récepteurs GABAA alpha1beta2gamma2 et alpha1beta2 recombinants. | Mercik, K., et al. 2006. Neuropharmacology. 51: 305-14. PMID: 16725161
- L'expression d'une mutation faux sens de la sorcine dans le cœur module le couplage excitation-contraction. | Collis, LP., et al. 2007. FASEB J. 21: 475-87. PMID: 17130302
- Les agonistes des récepteurs des benzodiazépines affectent à la fois la liaison et le passage des récepteurs recombinants alpha1beta2gamma2 de l'acide gamma-aminobutyrique. | Mercik, K., et al. 2007. Neuroreport. 18: 781-5. PMID: 17471066
- La ranolazine améliore la dysfonction diastolique dans le myocarde isolé de cœurs humains défaillants - rôle du courant sodique tardif et de l'accumulation d'ions intracellulaires. | Sossalla, S., et al. 2008. J Mol Cell Cardiol. 45: 32-43. PMID: 18439620
- Les endocannabinoïdes suppriment la transmission synaptique excitatrice aux neurones sérotoninergiques du raphé dorsal par l'activation des récepteurs CB1 présynaptiques. | Haj-Dahmane, S. and Shen, RY. 2009. J Pharmacol Exp Ther. 331: 186-96. PMID: 19592666
- La dysfonction diastolique et les arythmies causées par la surexpression de CaMKIIδ(C) peuvent être inversées par l'inhibition du courant Na(+) tardif. | Sossalla, S., et al. 2011. Basic Res Cardiol. 106: 263-72. PMID: 21174213
- Les effets inhibiteurs des analogues de l'oenanthotoxine sur les courants GABAergiques dans les neurones hippocampiques de rats en culture dépendent de la polarité des polyacétylènes. | Wyrembek, P., et al. 2012. Eur J Pharmacol. 683: 35-42. PMID: 22445880
- L'éthanol désinhibe les neurones épineux moyens du striatum dorsolatéral par l'activation d'un récepteur présynaptique delta opioïde. | Patton, MH., et al. 2016. Neuropsychopharmacology. 41: 1831-40. PMID: 26758662
- Le PIP2 dans les cellules β pancréatiques régule les canaux calciques gérés par le voltage par une voie indépendante du voltage. | de la Cruz, L., et al. 2016. Am J Physiol Cell Physiol. 311: C630-C640. PMID: 27488666
- Le stress en début de vie détermine les effets des glucocorticoïdes et du stress sur la fonction hippocampique: Preuves électrophysiologiques et comportementales respectivement. | Pillai, AG., et al. 2018. Neuropharmacology. 133: 307-318. PMID: 29412144
- Les cellules PDGFRβ relaient rapidement le signal inflammatoire du système circulatoire vers les neurones par l'intermédiaire de la chimiokine CCL2. | Duan, L., et al. 2018. Neuron. 100: 183-200.e8. PMID: 30269986
- L'épuisement des réserves de calcium intracellulaire par l'ionophore calcique A23187 induit les gènes des protéines régulées par le glucose dans les fibroblastes de hamster. | Drummond, IA., et al. 1987. J Biol Chem. 262: 12801-5. PMID: 3114264
- Les récepteurs potentiels transitoires TRPV1 et TRPA1 dans les mélanocytes sont responsables de la synergie entre la mélanogénèse dépendante et indépendante des UV. | Jia, Q., et al. 2021. Br J Pharmacol. 178: 4646-4662. PMID: 34363226
Informations pour la commande
| Nom du produit | Ref. Catalogue | COND. | Prix HT | QTÉ | Favoris | |
BAPTA-tetracesium Salt, 1 g | sc-503285 | 1 g | $398.00 |