")
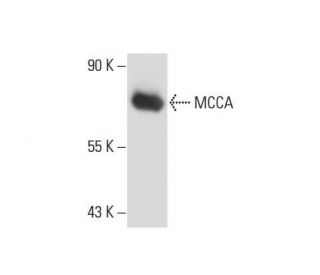
: sc-271427. Analyse par Western blot de l'expression de MCCA dans le lysat de cellules entières 3T3-L1.")

3T3-L1 Cell Lysate: sc-2243
- 500 µg protein in 200 µl SDS-PAGE Western blotting buffer
- mouse whole cell lysate; pre-adipose embryo fibroblast cells
- Lysat cellulaire total fourni comme contrôle positif en Western Blot
- should be stored at -20°C and repeated freezing and thawing should be minimized
- sample vial should be placed at 95° C for up to 5 minutes, once prior to use

ACCÈS RAPIDE AUX LIENS
VOIR ÉGALEMENT...
Le lysat cellulaire 3T3-L1 est un outil de recherche crucial dérivé de la lignée cellulaire 3T3-L1, qui provient de fibroblastes d'embryons de souris et est réputée pour sa capacité à se différencier en adipocytes dans des conditions appropriées. Ce lysat est obtenu par un processus minutieux de culture des cellules 3T3-L1, d'induction de l'adipogenèse, puis de lyse des cellules pour libérer leur contenu intracellulaire, qui comprend un large éventail de protéines, de lipides et d'autres composants cellulaires. Les chercheurs utilisent fréquemment le lysat de cellules 3T3-L1 pour étudier la biologie des adipocytes, en se concentrant particulièrement sur les mécanismes moléculaires qui sous-tendent l'adipogenèse, le métabolisme des lipides et les voies de signalisation de l'insuline. Ce lysat joue un rôle essentiel dans les études protéomiques et génomiques, en facilitant les analyses détaillées telles que le Western blotting, la spectrométrie de masse et le séquençage de l'ARN. En fournissant une riche source de protéines cellulaires et d'autres biomolécules, le lysat de cellules 3T3-L1 permet aux scientifiques d'étudier les effets de divers traitements et conditions sur la fonction et le métabolisme des adipocytes. En outre, il sert de référence précieuse pour la validation des anticorps et autres réactifs de recherche, contribuant ainsi à la reproductibilité et à la fiabilité des résultats expérimentaux. Dans l'ensemble, le lysat de cellules 3T3-L1 joue un rôle essentiel dans l'amélioration de notre compréhension de la physiologie des adipocytes et des processus métaboliques connexes dans le domaine de la recherche biologique.
3T3-L1 Cell Lysate Références:
- Les androgènes diminuent l'adiponectine plasmatique, une protéine dérivée des adipocytes qui sensibilise à l'insuline. | Nishizawa, H., et al. 2002. Diabetes. 51: 2734-41. PMID: 12196466
- Diminution de la production de leptine induite par l'oxyde nitrique dans les adipocytes 3T3-L1. | Unno, Y., et al. 2006. Nitric Oxide. 15: 125-32. PMID: 16442319
- Transfert aminoacyl d'un acide aminé non naturel à l'extrémité N-terminale de peptides et de protéines par la leucyl/phénylalanyl(L/F)-tRNA-protéine transférase et fonctionnalisation ultérieure par des réactions bioorthogonales. | Taki, M. and Sisido, M. 2007. Biopolymers. 88: 263-71. PMID: 17216634
- L'autorisation et l'exécution de l'adipogenèse sont liées de manière disparate à la prolifération cellulaire. | Guo, W., et al. 2009. Cell Res. 19: 216-23. PMID: 19065151
- De nouveaux messages polysomiques et des changements dans l'activité de traduction apparaissent après l'induction de l'adipogenèse dans les cellules 3T3-L1. | Fromm-Dornieden, C., et al. 2012. BMC Mol Biol. 13: 9. PMID: 22436005
- Propriétés anti-obésité d'un extrait de Sasa quelpaertensis chez des souris obèses induites par un régime riche en graisses. | Kang, SI., et al. 2012. Biosci Biotechnol Biochem. 76: 755-61. PMID: 22484945
- La fisétine augmente l'expression de l'adiponectine dans les adipocytes 3T3-L1 via l'activation de la désacétylase SIRT1 (silent mating type information regulation 2 homologue 1) et des récepteurs activés par les proliférateurs de peroxysomes (PPAR). | Jin, T., et al. 2014. J Agric Food Chem. 62: 10468-74. PMID: 25286082
- Une stratégie alternative pour la génération d'anticorps à la pan-acétyl-lysine. | Kim, SY., et al. 2016. PLoS One. 11: e0162528. PMID: 27606599
- L'activité hypolipidémique de l'extrait d'éthanol du fruit de Quercus acutissima est médiée par l'inhibition de l'acétylation. | Hwang, JT., et al. 2017. J Med Food. 20: 542-549. PMID: 28581876
- La désactivation de l'aquaporine-8 induit un dysfonctionnement mitochondrial dans les cellules 3T3-L1. | Ikaga, R., et al. 2015. Biochem Biophys Rep. 4: 187-195. PMID: 29124204
- L'exposition des adipocytes au bisphénol-A in vitro interfère avec l'action de l'insuline sans favoriser l'adipogenèse. | De Filippis, E., et al. 2018. PLoS One. 13: e0201122. PMID: 30133442
- La lipolyse médiée par Nod1 favorise l'accumulation de diacylglycérol et l'inflammation successive via l'axe PKCδ-IRAK dans les adipocytes. | Sharma, A., et al. 2019. Biochim Biophys Acta Mol Basis Dis. 1865: 136-146. PMID: 30391544
- Développement et validation d'une méthode LC-MS/MS pour la tyrphostine A9. | Meyer, LF. and Shah, DK. 2019. J Pharm Anal. 9: 163-169. PMID: 31297293
- KCTD1: Un nouveau modulateur de l'adipogenèse par l'interaction avec le facteur de transcription AP2α. | Pirone, L., et al. 2019. Biochim Biophys Acta Mol Cell Biol Lipids. 1864: 158514. PMID: 31465887
- L'apigénine inhibe l'axe de signalisation STAT3/CD36 et réduit l'obésité viscérale. | Su, T., et al. 2020. Pharmacol Res. 152: 104586. PMID: 31877350
Informations pour la commande
| Nom du produit | Ref. Catalogue | COND. | Prix HT | QTÉ | Favoris | |
3T3-L1 Cell Lysate | sc-2243 | 500 µg/200 µl | $120.00 |